


Next: 2 Environmental Effects
Up: 4 The Elements of
Previous: 4 The Elements of
Index
A complete understanding of genetic effects would need to answer
a series of questions:
- How important are genetic effects on human differences?
- What kinds of action and interaction occur between gene products in
the pathways between genotype and phenotype?
- Are the genetic effects on a trait consistent across sexes?
- Are there some genes that have particularly outstanding effects when
compared to others?
- Whereabouts on the human gene map are these genes located?
Questions 4 and 5 are clearly very important, but are not the
immediate concern of this book. On the other hand, we shall have a
lot to say about 1, 2, and 3. It is arguable that we shall not be
able to understand 4 and 5 adequately, if we do not have a proper
appreciation of these other issues.
The importance of genes is often expressed relative to all
the causes of variation, both genetic and environmental. The
proportion of variation associated with genetic effects is termed the
broad heritability. However, the
complete analysis of genetic factors does not end here because, as
countless experiments in plant and animal genetics have shown (well in
advance of modern molecular genetics; see e.g., Mather and Jinks,
1982), genes can act and interact in a variety of ways before their
effects on the phenotype appear.
Geneticists typically distinguish between additive and
non-additive genetic effects (these terms will be defined more
explicitly in Chapter 3). These influences have been studied
in
detail in many non-human species using selective breeding
experiments, which directly alter the frequencies of particular
genotypes. In such experiments, the bulk of genetic variation is
usually explained by additive genetic effects.
However, careful studies have shown two general types of
non-additivity that may be important, especially in traits that have
been subject to strong directional selection in the wild. The two
main types of genetic non-additivity are dominance and
epistasis.
The term dominance derives initially from Mendel's classical
experiments in which it was shown that the progeny of a cross between
two pure breeding lines often resembled one parent more than the
other. That is, an individual who carries different alleles at a
locus (the heterozygote) is not
exactly intermediate in expression between
individuals who are pure breeding (homozygous) for the two
alleles. While dominance describes the interaction between alleles at
the same locus, epistasis describes the interaction between
alleles at different loci.
Epistasis is said to occur whenever the effects of one gene on
individual differences depend on which genotype is expressed at
another locus. For example, suppose that at locus 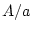 individuals
may have genotype
individuals
may have genotype 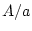 ,
, 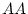 or
or 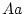 , and at locus
, and at locus 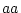 genotype
genotype
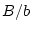 ,
, 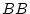 or
or 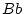
![[*]](footnote.png) . If the difference between individuals with
genotype and those with genotype depends on whether they are
or , then there would be additive
. If the difference between individuals with
genotype and those with genotype depends on whether they are
or , then there would be additive  additive epistatic interactions. Experimental studies have shown a
rich variety of possible epistatic interactions depending on the
number and effects of the interacting loci. However, their detailed
resolution in humans is virtually impossible unless we are fortunate
enough to be examining a trait which is influenced by a small number
of known genetic loci. Therefore we acknowledge their conceptual
importance and model them if they are identified. Failure to take
non-additive genetic effects into account may be one of the main
reasons studies of twins give different heritability estimates from
studies of adoptees and nuclear families (Eaves
et al., 1992; Plomin et al.,
1991).
As studies in genetic epidemiology become larger and better
designed, it is becoming increasingly clear that there are marked
sex differences in gene expression. An important factor in
establishing this view has been the incorporation of unlike-sex twin
pairs in twin studies (Eaves et al., 1990).
However, comparison of statistics derived from any relationship of
individuals of unlike sex with those of like sex would yield a similar
conclusion (see Chapter 9). We shall make an important
distinction between two types of sex-limited gene expression. In
the simpler case,
the
same genes affect both males and females, but their effects are
consistent across sexes and differ only by some constant multiple over
all the loci involved. We shall refer to this type of effect as
scalar sex-limitation. In other
cases, however, we shall discover that genetic effects in one sex are
not just a constant multiple of their effects in the other. Indeed,
even though a trait may be measured in exactly the same way in males
and females, it may turn out that quite different genes control its
expression in the two sexes. A classic example would be the case of
chest-girth since at least some of the variation expressed in females
may be due to loci that, while still present in males, are only
expressed in females. In this case we shall speak of non-scalar
sex-limitation. None of us
likes the term very much, but until someone suggests something better
we shall continue to use it!
additive epistatic interactions. Experimental studies have shown a
rich variety of possible epistatic interactions depending on the
number and effects of the interacting loci. However, their detailed
resolution in humans is virtually impossible unless we are fortunate
enough to be examining a trait which is influenced by a small number
of known genetic loci. Therefore we acknowledge their conceptual
importance and model them if they are identified. Failure to take
non-additive genetic effects into account may be one of the main
reasons studies of twins give different heritability estimates from
studies of adoptees and nuclear families (Eaves
et al., 1992; Plomin et al.,
1991).
As studies in genetic epidemiology become larger and better
designed, it is becoming increasingly clear that there are marked
sex differences in gene expression. An important factor in
establishing this view has been the incorporation of unlike-sex twin
pairs in twin studies (Eaves et al., 1990).
However, comparison of statistics derived from any relationship of
individuals of unlike sex with those of like sex would yield a similar
conclusion (see Chapter 9). We shall make an important
distinction between two types of sex-limited gene expression. In
the simpler case,
the
same genes affect both males and females, but their effects are
consistent across sexes and differ only by some constant multiple over
all the loci involved. We shall refer to this type of effect as
scalar sex-limitation. In other
cases, however, we shall discover that genetic effects in one sex are
not just a constant multiple of their effects in the other. Indeed,
even though a trait may be measured in exactly the same way in males
and females, it may turn out that quite different genes control its
expression in the two sexes. A classic example would be the case of
chest-girth since at least some of the variation expressed in females
may be due to loci that, while still present in males, are only
expressed in females. In this case we shall speak of non-scalar
sex-limitation. None of us
likes the term very much, but until someone suggests something better
we shall continue to use it!
Next: 2 Environmental Effects
Up: 4 The Elements of
Previous: 4 The Elements of
Index
Jeff Lessem
2002-03-21