We illustrate the estimation of the genetic and environmental
covariance matrices for a simple case of skinfold measures
made on 11.5 year-old male twins from the
Medical College of Virginia Twin Study
(Schieken et al., 1989)![[*]](footnote.png) Our skinfold assessments include
four different measures which were obtained using standard
anthropometric techniques. The measures were obtained for biceps,
subscapular, suprailiac, and triceps skinfolds. The raw data were
averaged for the left and right sides and subjected to a logarithmic
transformation prior to analysis in order to remove the correlation
between error variance and skinfold measure. The
Our skinfold assessments include
four different measures which were obtained using standard
anthropometric techniques. The measures were obtained for biceps,
subscapular, suprailiac, and triceps skinfolds. The raw data were
averaged for the left and right sides and subjected to a logarithmic
transformation prior to analysis in order to remove the correlation
between error variance and skinfold measure. The 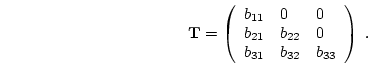 covariance matrices for the male MZ and DZ twins are given in
Table 10.3.
covariance matrices for the male MZ and DZ twins are given in
Table 10.3.
| Dizygotic Male Pairs (N=33) |
||||||||
| BIC1 | SSC1 | SUP1 | TRI1 | BIC2 | SSC2 | SUP2 | TRI2 | |
| BIC1 | .1538 | |||||||
| SSC1 | .1999 | .3007 | ||||||
| SUP1 | .2266 | .3298 | .3795 | |||||
| TRI1 | .1285 | .1739 | .2007 | .1271 | ||||
| BIC2 | .0435 | .0336 | .0354 | .0376 | .1782 | |||
| SSC2 | .0646 | .0817 | .0741 | .0543 | .2095 | .3081 | ||
| SUP2 | .0812 | .0901 | .0972 | .0666 | .2334 | .3241 | .3899 | |
| TRI2 | .0431 | .0388 | .0376 | .0373 | .1437 | .1842 | .2108 | .1415 |
| Monozygotic Male Pairs (N=84) |
||||||||
| BIC1 | SSC1 | SUP1 | TRI1 | BIC2 | SSC2 | SUP2 | TRI2 | |
| BIC1 | .1285 | |||||||
| SSC1 | .1270 | .1759 | ||||||
| SUP1 | .1704 | .2156 | .3031 | |||||
| TRI1 | .1035 | .1101 | .1469 | .1041 | ||||
| BIC2 | .0982 | .1069 | .1491 | .0824 | .1233 | |||
| SSC2 | .0999 | .1411 | .1848 | .0880 | .1295 | .1894 | ||
| SUP2 | .1256 | .1654 | .2417 | .1095 | .1616 | .2185 | .2842 | |
| TRI2 | .0836 | .0907 | .1341 | .0836 | .1010 | .1134 | .1436 | .1068 |
| Variable Labels: BIC=Biceps; SSC=Subscapular; SUP=Suprailiac; | ||||||||
| TRI=Triceps. ``1'' and ``2'' refer to measures on first and second twins | ||||||||
An example Mx program for estimating the Cholesky factors of the
additive genetic and within-family environmental covariance matrices
is given in Appendix ![[*]](crossref.png) . The matrices X and Z are now declared as free lower triangular matrices.
. The matrices X and Z are now declared as free lower triangular matrices.
When this program is run with the data from male twins, we obtain a
goodness-of-fit chi-squared of 68.92 for 52 d.f. (![]() )
suggesting that the AE model gives a reasonable fit to these data.
Setting the off-diagonal elements of the genetic factors to zero
yields a chi-squared that may be compared using the difference test to
see whether the measures can be regarded as genetically independent.
This chi-squared turns out to be 110.96 for 6 d.f. which is highly
significant. Therefore, the genetic correlations between these
skinfold measures cannot be ignored. Similarly, setting the
environmental covariances to zero yields a significant increase in
chi-squared of 356.98, also for 6 d.f. Clearly, there are also highly
significant environmental covariances among the four variables.
)
suggesting that the AE model gives a reasonable fit to these data.
Setting the off-diagonal elements of the genetic factors to zero
yields a chi-squared that may be compared using the difference test to
see whether the measures can be regarded as genetically independent.
This chi-squared turns out to be 110.96 for 6 d.f. which is highly
significant. Therefore, the genetic correlations between these
skinfold measures cannot be ignored. Similarly, setting the
environmental covariances to zero yields a significant increase in
chi-squared of 356.98, also for 6 d.f. Clearly, there are also highly
significant environmental covariances among the four variables.
Table 10.4 gives the estimates of the Cholesky factors of
the genetic and environmental covariance matrices produced by Mx.
| Genetic Factor | Environmental Factor | ||||||||
| Variable | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | |
| Biceps | 0.340 | 0.000 | 0.000 | 0.000 | 0.170 | 0.000 | 0.000 | 0.000 | |
| Subscapular | 0.396 | 0.182 | 0.000 | 0.000 | 0.160 | 0.138 | 0.000 | 0.000 | |
| Suprailiac | 0.487 | 0.159 | 0.148 | 0.000 | 0.180 | 0.117 | 0.093 | 0.000 | |
| Triceps | 0.288 | 0.016 | 0.036 | 0.110 | 0.117 | 0.039 | -0.004 | 0.085 | |
| Genetic | Environmental | ||||||||
| Variable | BIC | SSC | SUP | TRI | BIC | SSC | SUP | TRI | |
| Biceps | 0.116 | 0.135 | 0.166 | 0.098 | 0.029 | 0.027 | 0.030 | 0.020 | |
| Subscapular | 0.909 | 0.190 | 0.222 | 0.117 | 0.759 | 0.044 | 0.045 | 0.024 | |
| Suprailiac | 0.914 | 0.955 | 0.284 | 0.148 | 0.769 | 0.908 | 0.054 | 0.025 | |
| Triceps | 0.927 | 0.863 | 0.894 | 0.097 | 0.778 | 0.757 | 0.716 | 0.023 | |
| Note: The variances are given on the diagonals of the two matrices; | |||||||||
We see that the genetic correlations between the four skin-fold measures are indeed very large, suggesting that the amount of fat at different sites of the body is almost entirely under the control of the same genetic factors. However, in this example, the environmental correlations also are quite large, suggesting that environmental factors which affect the amount of fat at one site also have a generalized effect over all sites.